MIT molecular Biology 笔记10 翻译
视频 https://www.bilibili.com/video/av7973580/
教材 Molecular biology of the gene 7th edition J.D. Watson et. al
翻译
翻译机器的组成
- mRNA
- tRNA
- aminoacyl tRNA synthetase
- ribosome
一、信使RNA
1、多肽链是由可读框决定的
- 含有多个ORF的mRNA叫多顺反子
- 只有1个ORF的mRNA叫单顺反子
2、原核细胞mRNA具有核糖体结合位点,可以募集翻译机器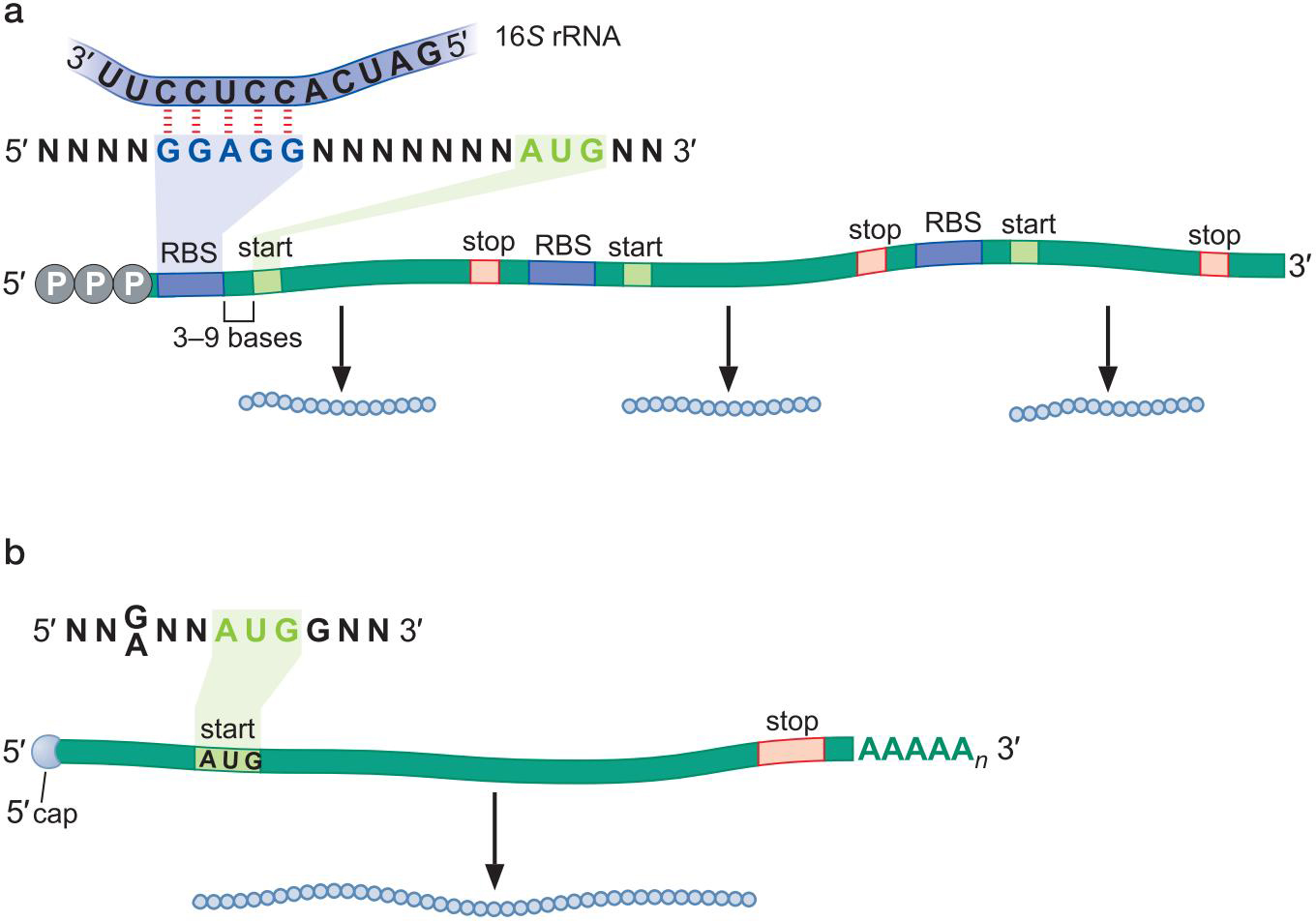
- 核糖体结合位点(RBS)
- 一般位于
密码子5'端 3~9bp内
- 与核糖体 16S rRNA 的 3'端互补(5' CCUCCU 3')
- 某些原核细胞ORF缺乏RBS也能翻译,是因为ORF紧接着上一个ORF:翻译耦合translational coupling
3、真核细胞mRNA在5' 和 3' 被修饰,促进翻译
- 真核mRNA的 5'帽是甲基化鸟嘌呤核苷酸
- 可以募集核糖体结合
- 核糖体向下游扫描,寻找AUG
- 在某些mRNA中,起始密码子周围有些保守位点
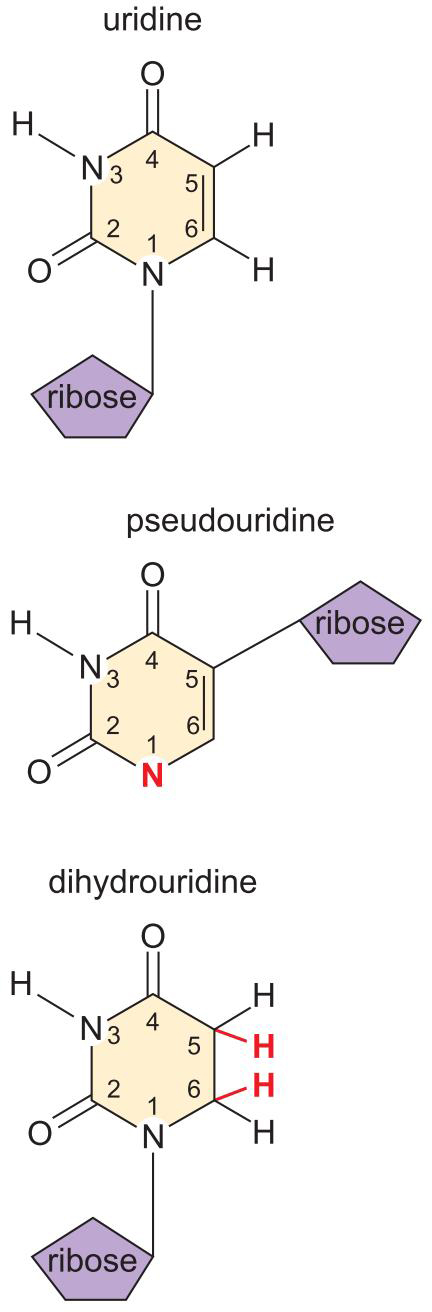
- 5'- G/ANNAUGG-3' Kozak序列:与起始tRNA互作
- 3' poly-A 尾: 增强5'Cap 的募集作用
- 帽和尾 稳定RNA
二、转运RNA
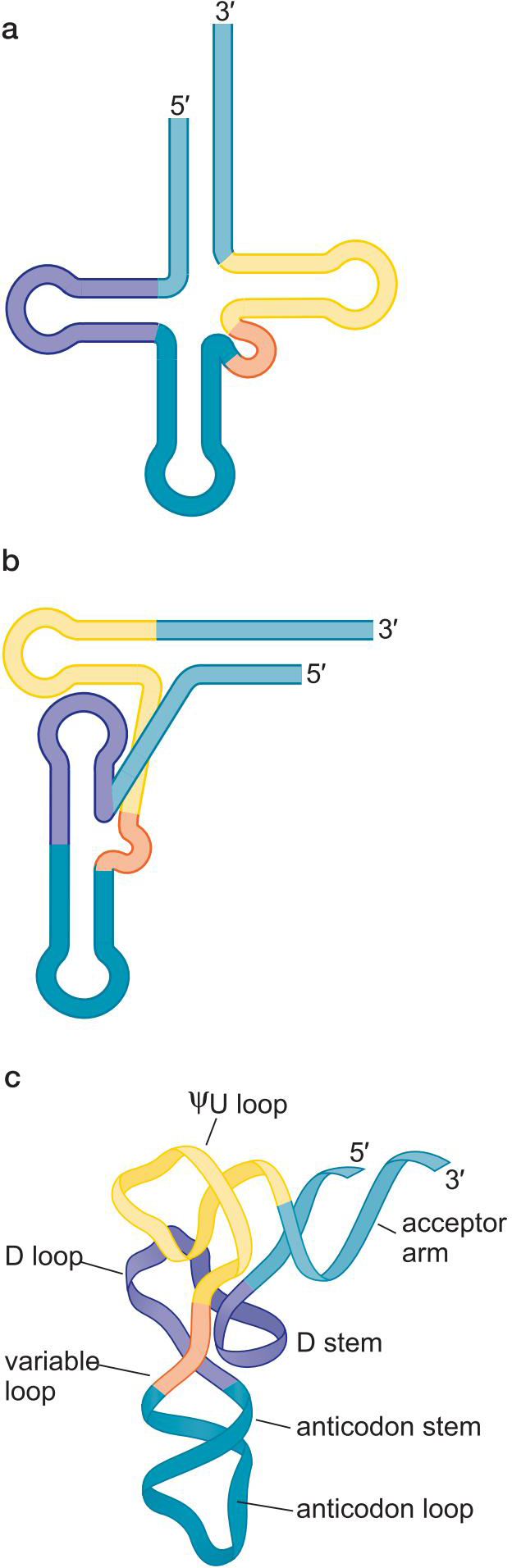
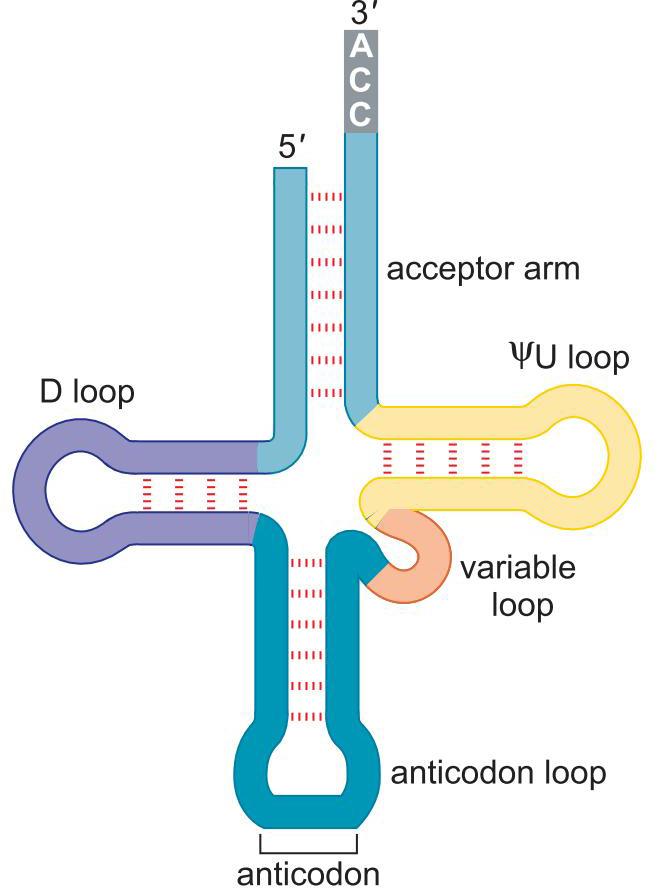
1、tRNA是密码子与氨基酸之间的转配器
特征
- 3'端均为5'-CCA-3' :氨基酸结合位点
- 存在特殊碱基:
- ΨU 假尿嘧啶
- D 双氢尿嘧啶
- H 次黄嘌呤
- T胸腺嘧啶
- Gm 甲基鸟嘌呤
2、共有三叶草二级结构
- 受体臂
- ΨU loop
- D loop
- anticodon loop
- variable loop
3、tRNA具有L形三维结构
三、氨基酸连接到tRNA上
1、氨基酸通过高能酰基连接到tRNA 3'端腺苷酸上使tRNA负载
- 腺苷酰基化 adenylylation(ATP)
- tRNA负载
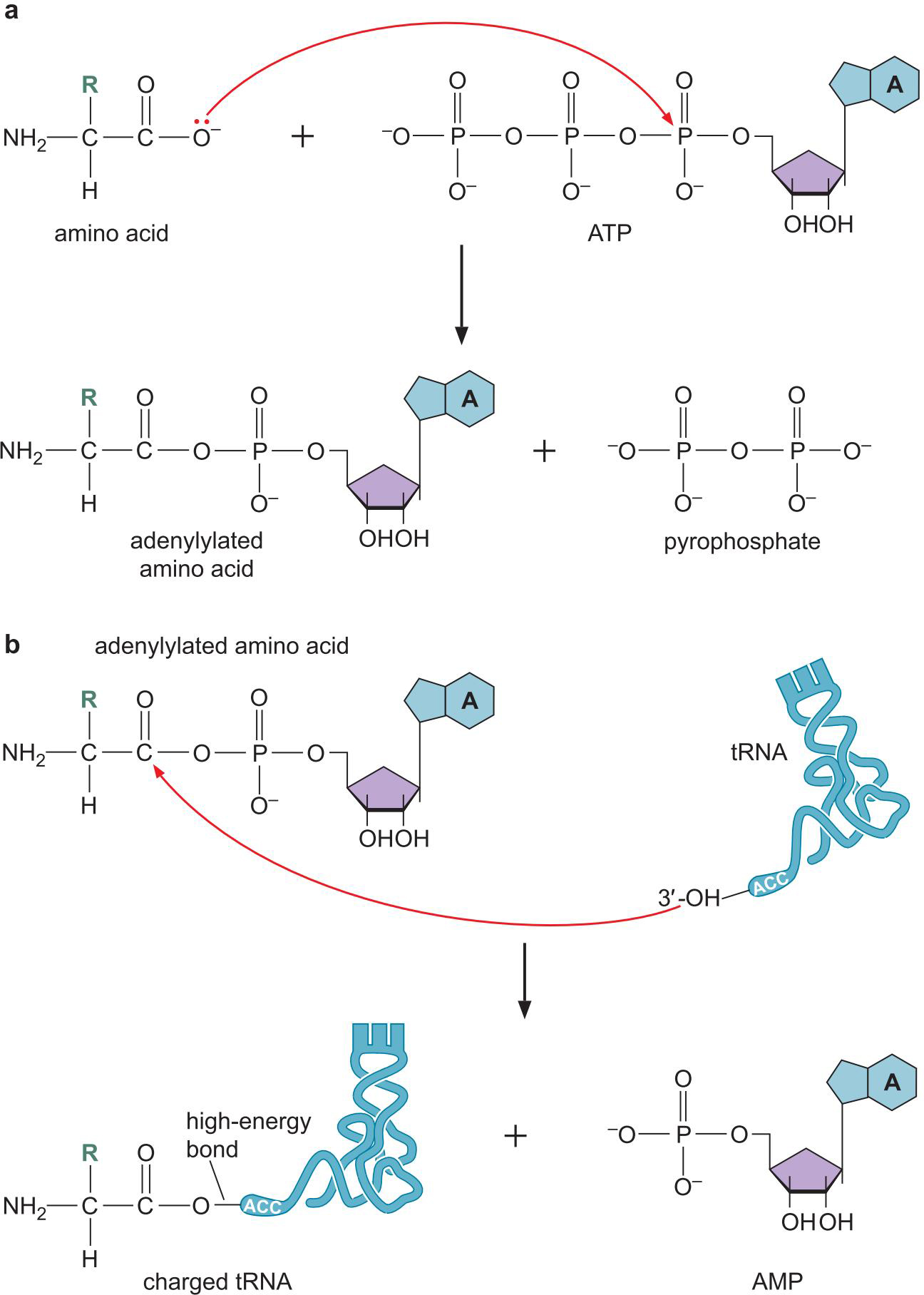
2、每种氨酰-tRNA合成酶连接一种氨基酸到一种或多种tRNA上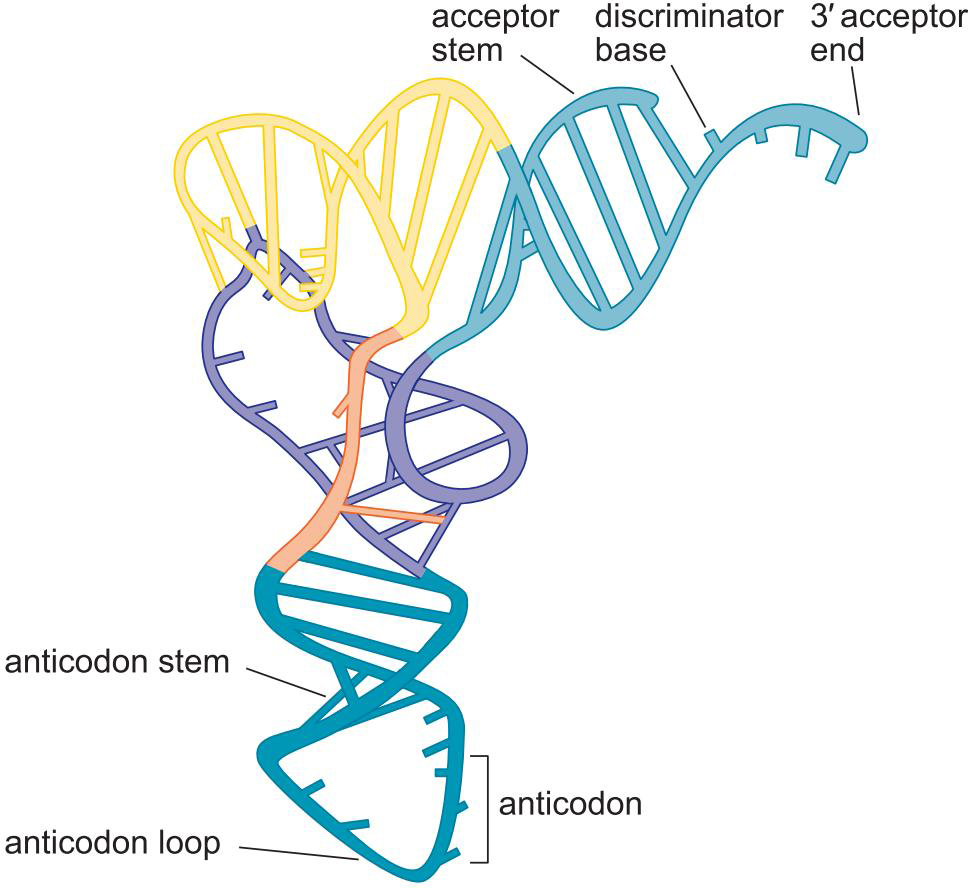
每个氨基酸对应一种合成酶
- 特殊的合成方式:Glu合成到tRNAGln,Glu在氧化成Gln
3、tRNA合成酶识别位于同族(cognate)tRNA上独特结构特征
- 受体臂上鉴别者碱基
- 反密码子环
4、保证氨酰-tRNA合成的精确
- 酶口袋的尺寸阻止大氨基酸进入
- 酶中的编辑口袋水解尺寸过小的氨基酸
- 那么只有正确的氨基酸能合成
5、核糖体不能辨别tRNA的负载是否正确
6、硒代半胱氨酸
- 存在于谷胱甘肽过氧化物酶,甲酸盐脱氢酶
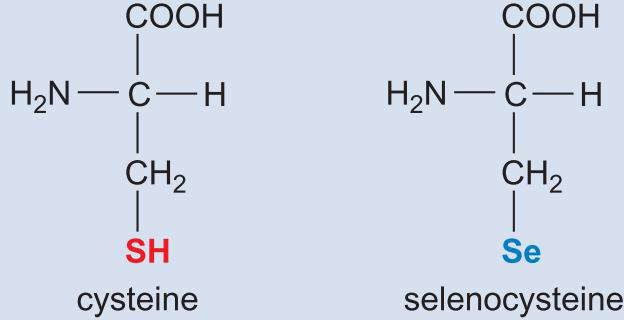
- 由UGA终止密码合成:特殊地机制
四、核糖体
- DNA复制速度 200~100bp/s
- 转录速度 50~100bp/s
- 翻译 原核 20AA/s 真核 2~4AA/s
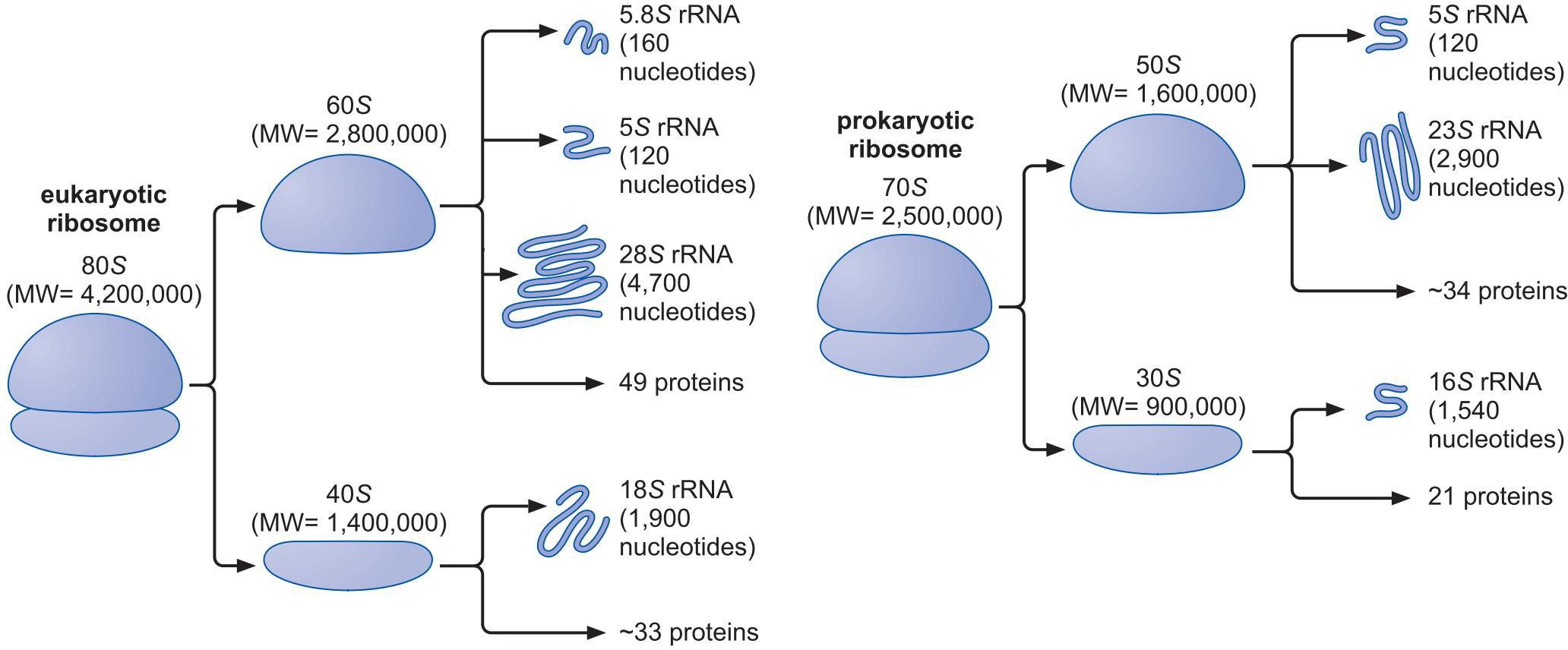
1、核糖体由一个大亚基,一个小亚基组成
- 大亚基:肽基转移酶中心 peptidyl transferase center
- 小亚基:解码中心 decoding center
- 大小亚基按照沉降速录命名(不和质量成严格正比)
2、核糖体循环

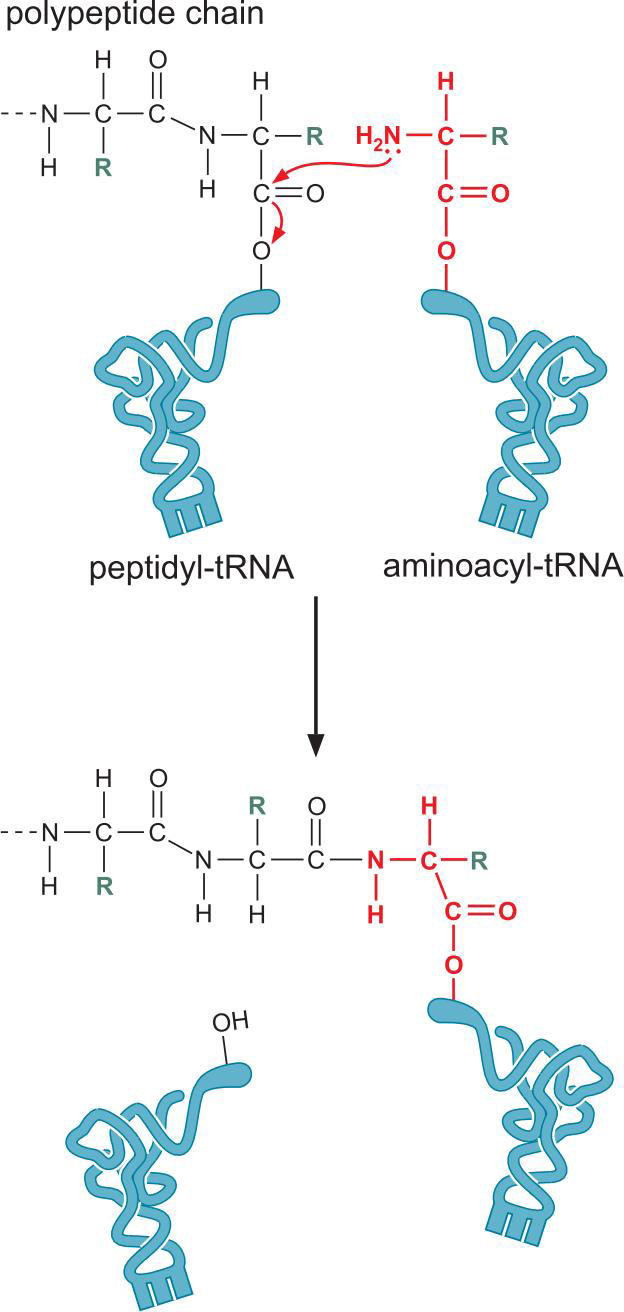
- 合成方向 N->C
- 形成肽键的过程:将正在延伸的多肽转移到另一个tRNA上
- 肽键的形成不需NTP水解:氨酰-tRNA储存着能量
3、核糖体RNA是核糖体中结构和催化功能决定因子
- 以RNA为中心
4、核糖体有3个tRNA结合位点
- A 氨酰tRNA结合
- P 肽酰tRNA结合
- E tRNA离开位点
5、核糖体内的通道使mRNA与延伸多肽进出
五、翻译的起始
- 核糖体募集
- 负载tRNA置于核糖体P位点
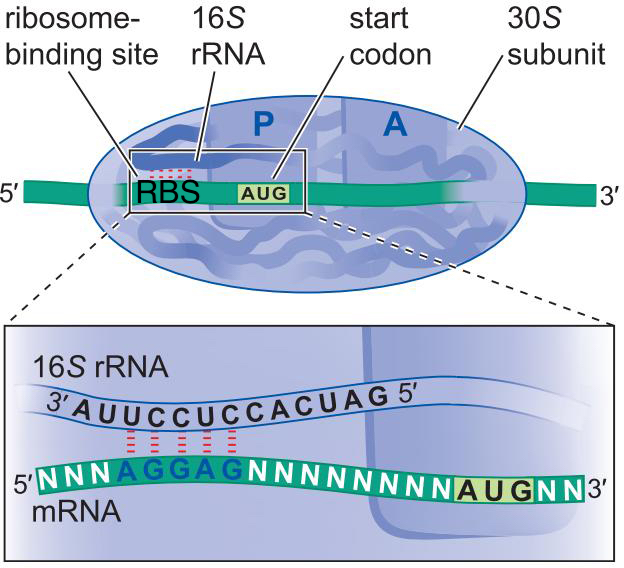
- 核糖体精确定位
1、原核细胞
- 三种起始因子指导 mRNA 和 tRNA 装配起始复合物
- IF1 结合到A位点,防止起始tRNA结合
- IF2 GTP酶,促进起始fMet-tRNAifMet结合至P位点,阻止别的结合
- IF3 与小亚基结合,阻止翻译起始前大亚基结合(最后结合)
- 随着起始因子的加入,小亚基准备好和mRNA,tRNA以任意顺序结合
- 与mRNA结合:RBS 与 16S RNA相互作用
- 结合后,起始密码子正好处于P位点
- 起始 fMet-tRNAifMet 的结合:通过GTP与IF2相互作用
- 起始的Met被加上甲酰基(Met-tRNA转甲酰酶) :f-N -> C
- 起始密码子与起始tRNA配对:小亚基构象变化,IF3释放
- 大亚基结合(IF2作为初始对接位点),形成70S 起始复合体
- IF2、IF1 释放
- 许多蛋白质不是以Met开始的,氨肽酶通常会在氨基酸切除Met及另外一两个氨基酸
2、真核细胞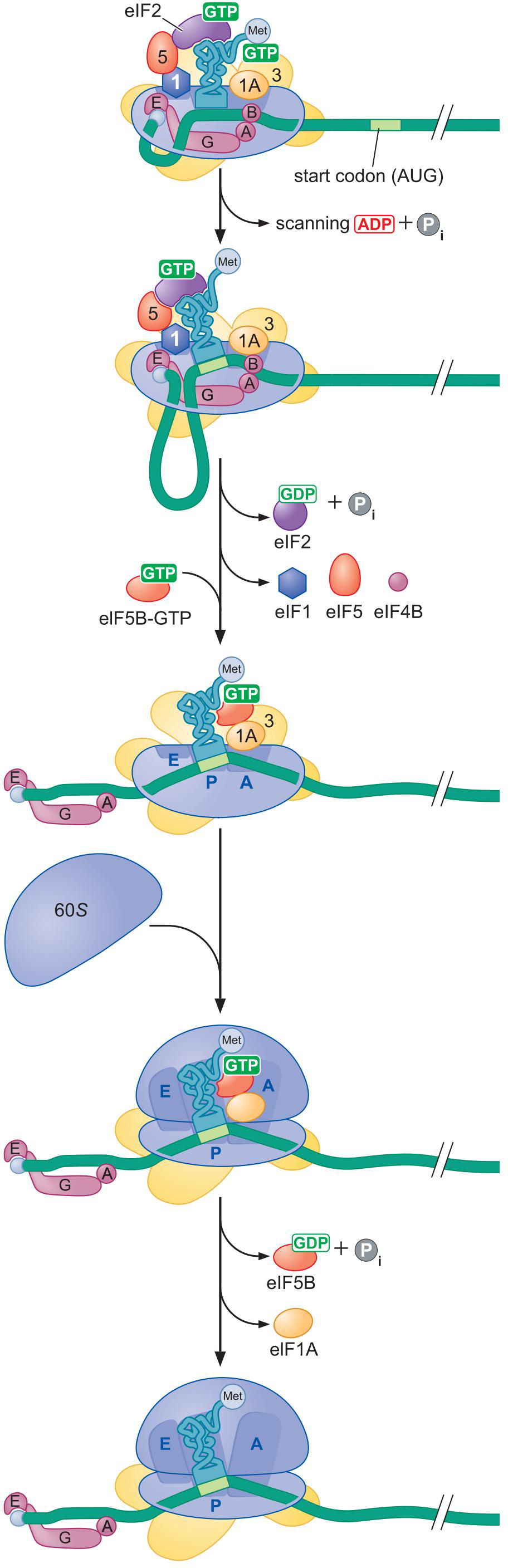
- 真核细胞中tRNA结合先于mRNA的结合
- 使用另一组辅助因子
- 核糖体小亚基结合上mRNA 的 5'Cap,扫描AUG(ATP)
- 扫描到AUG后,组装大亚基,开始翻译
- 翻译起始因子使真核mRNA保持环状:提高翻译效率
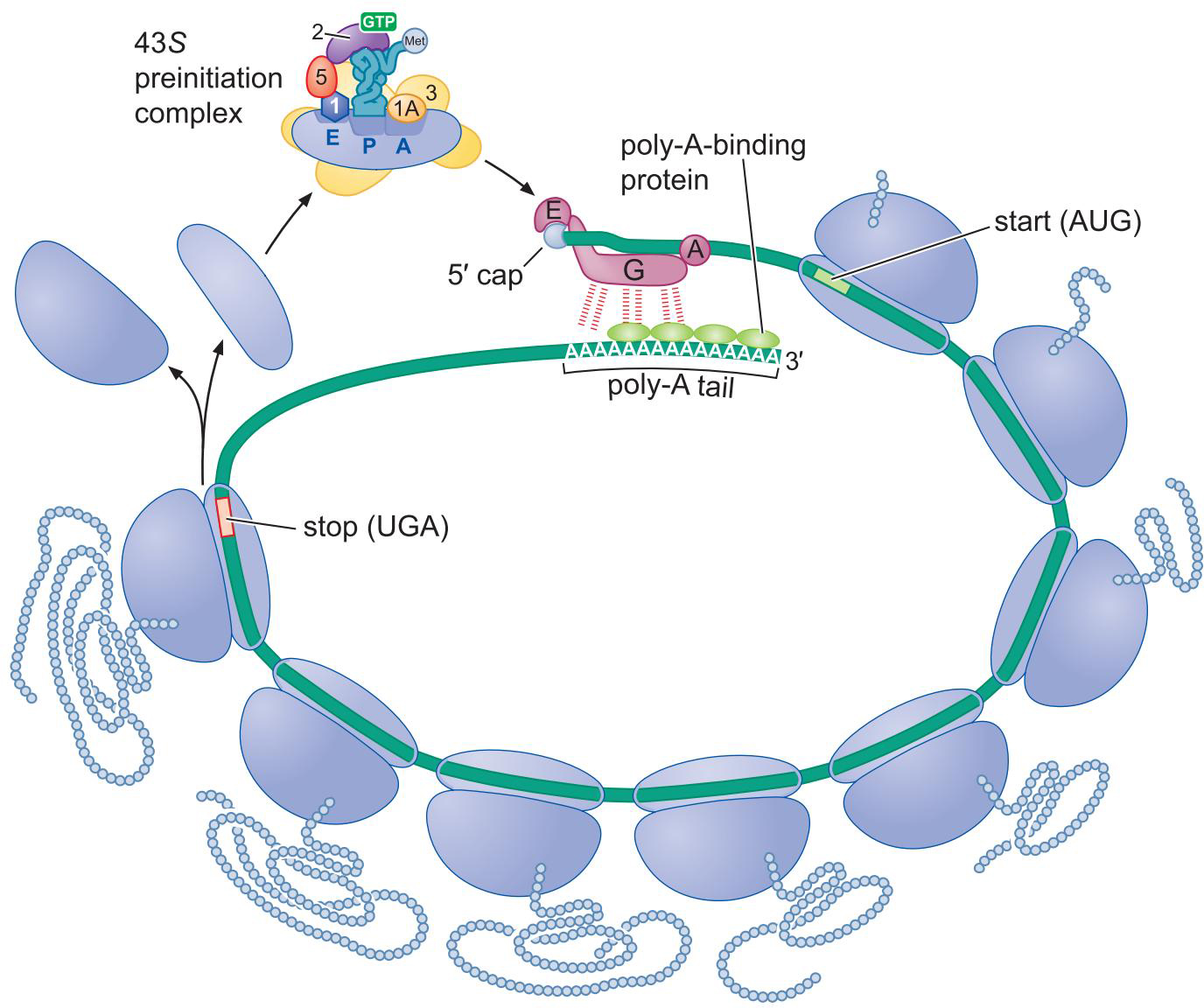
六、翻译的延伸
- 氨酰-tRNA结合上A位点:EF-Tu-GTP辅助
- 延伸:肽酰转移酶将多肽链加到A位点负载tRNA上
- 易位
- 在两种延伸因子的辅助下,正确的阅读ORF
氨酰tRNA结合
1、氨酰-tRNA由延伸因子EF-Tu送达A位点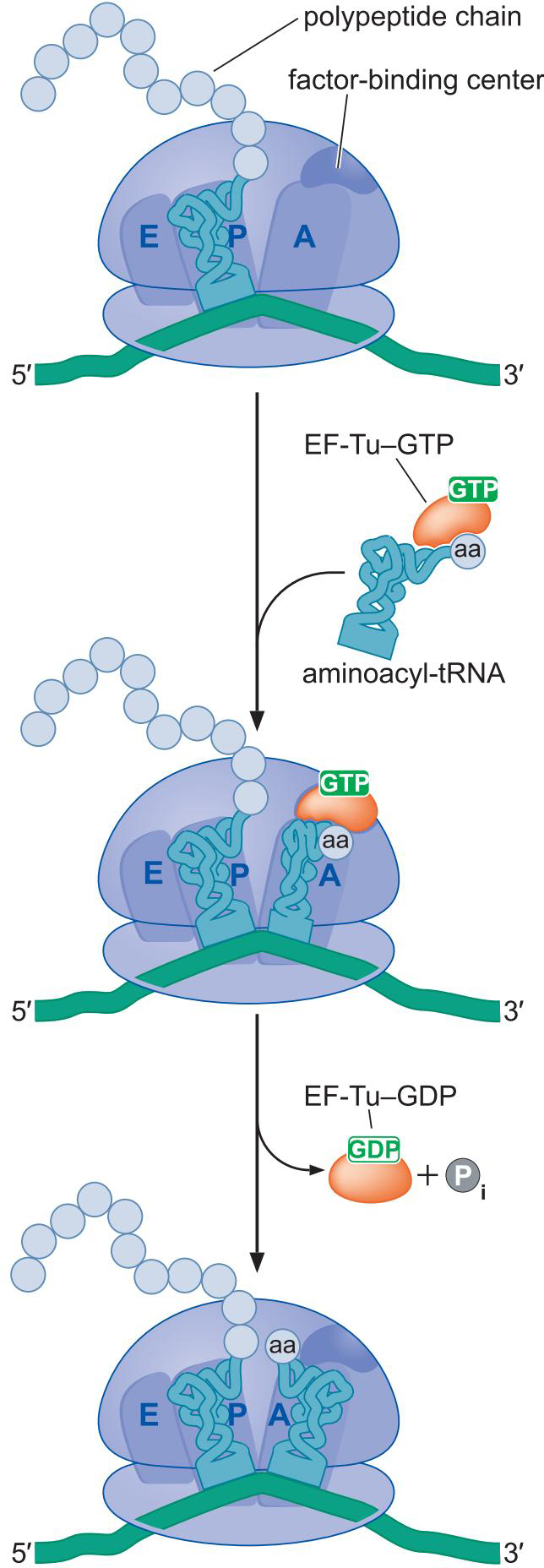
- EF-Tu-GTP覆盖tRNA3'端的氨基酸:防止过早反应
- 正确匹配结合上A位点后,大亚基的某结构域激活GTP水解
- EF-Tu-GDP释放
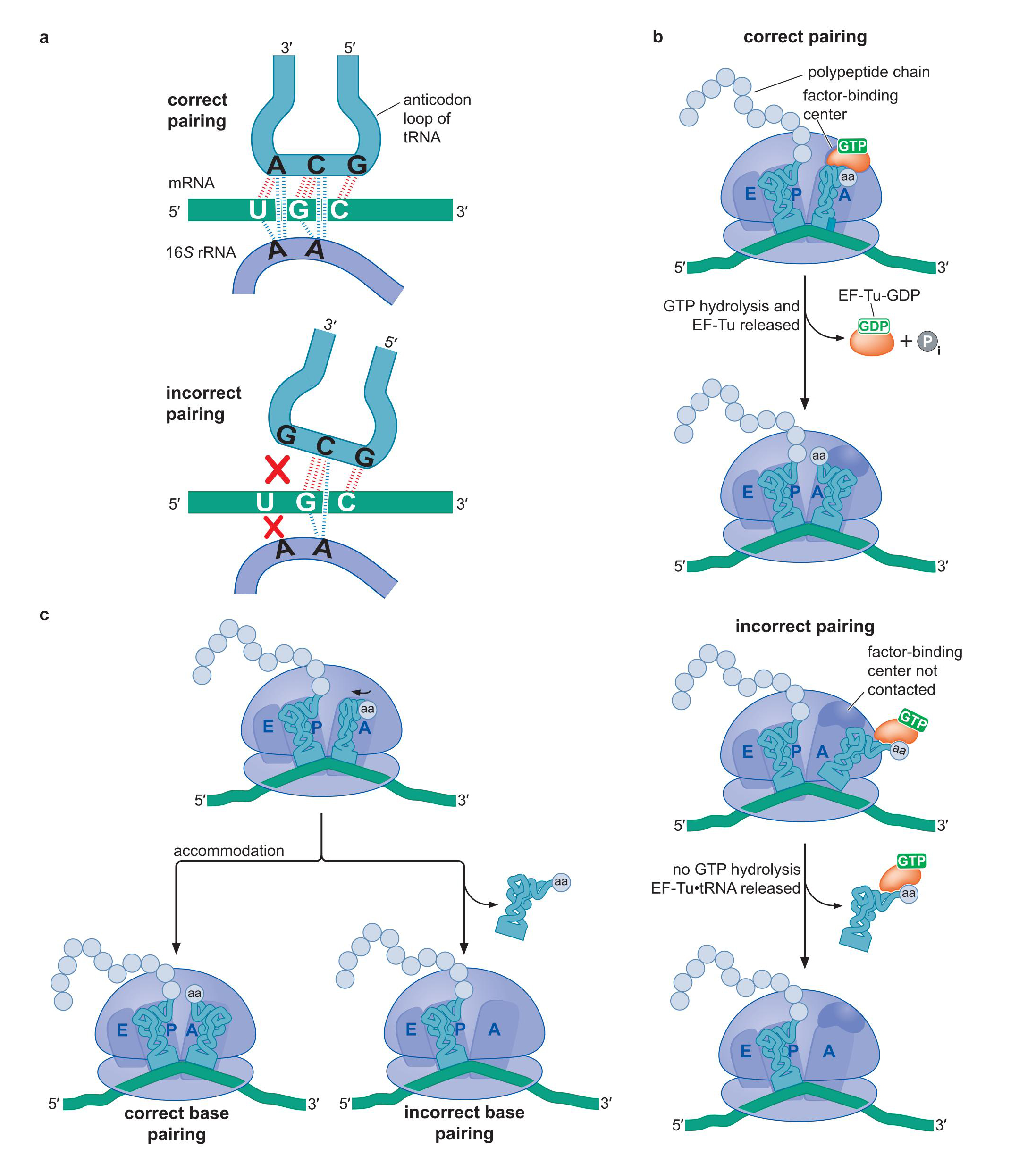
2、核糖体利用多种机制排斥错误配对的 氨酰-tRNA
- 小亚基 16S RNA的两个碱基识别 tRNA-mRNA 之间的小沟:错配小沟形状不符合
- EF-Tu-GTP=tRNA复合物要处于正确位点,才能导致GTP水解
- 易位时,错误配对的tRNA容易脱落
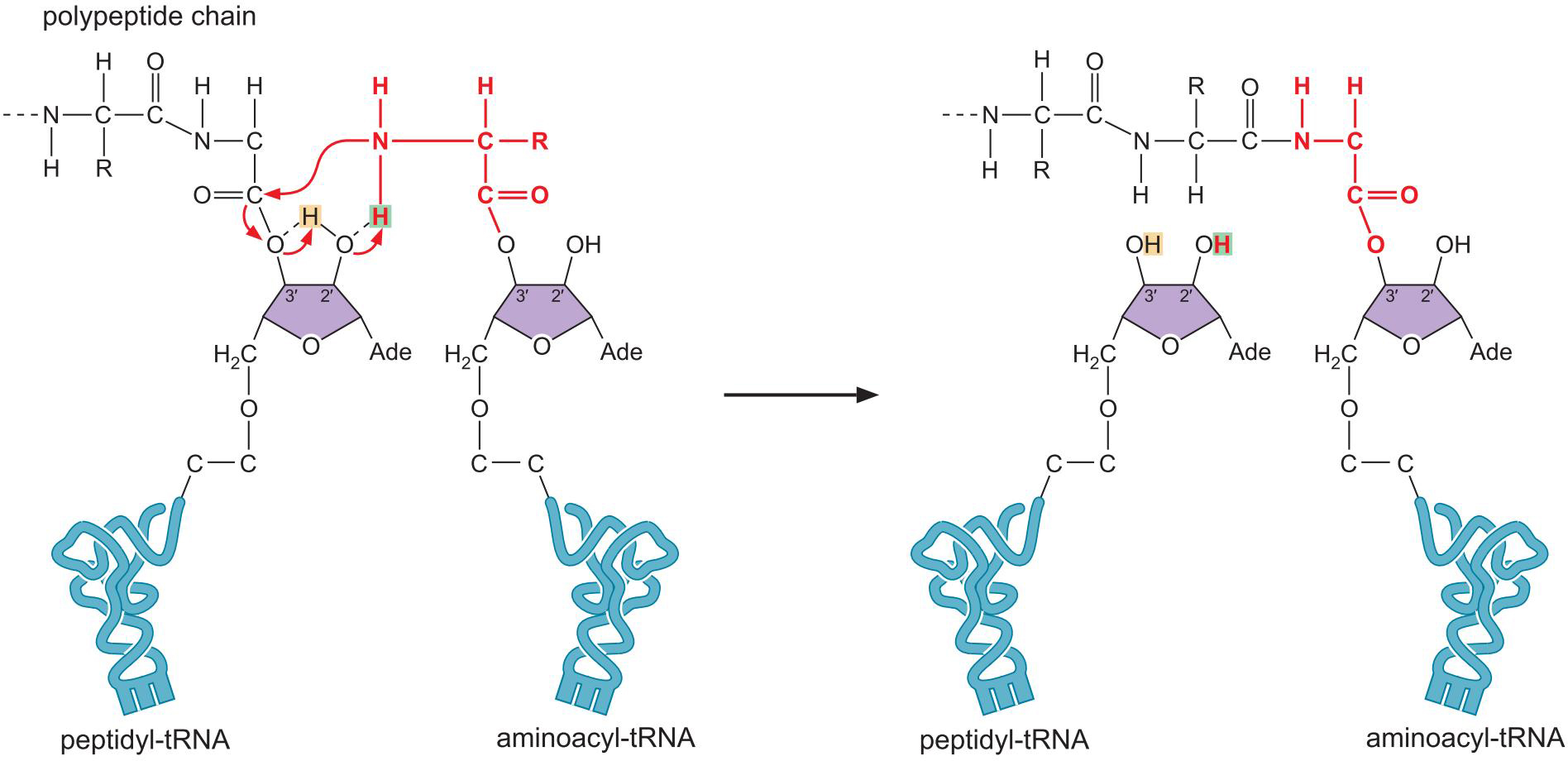
肽键的形成
3、核糖体是一种核酶
- rRNA处于活性中心,蛋白起辅助作用
- 熵催化 entropic catalysis
- rRNA通过碱基配对,将底物稳定在活性中心已促进反应
- 底物辅助催化
- P 位点 tRNA 的 2'OH 通过质子穿梭机制辅助催化
易位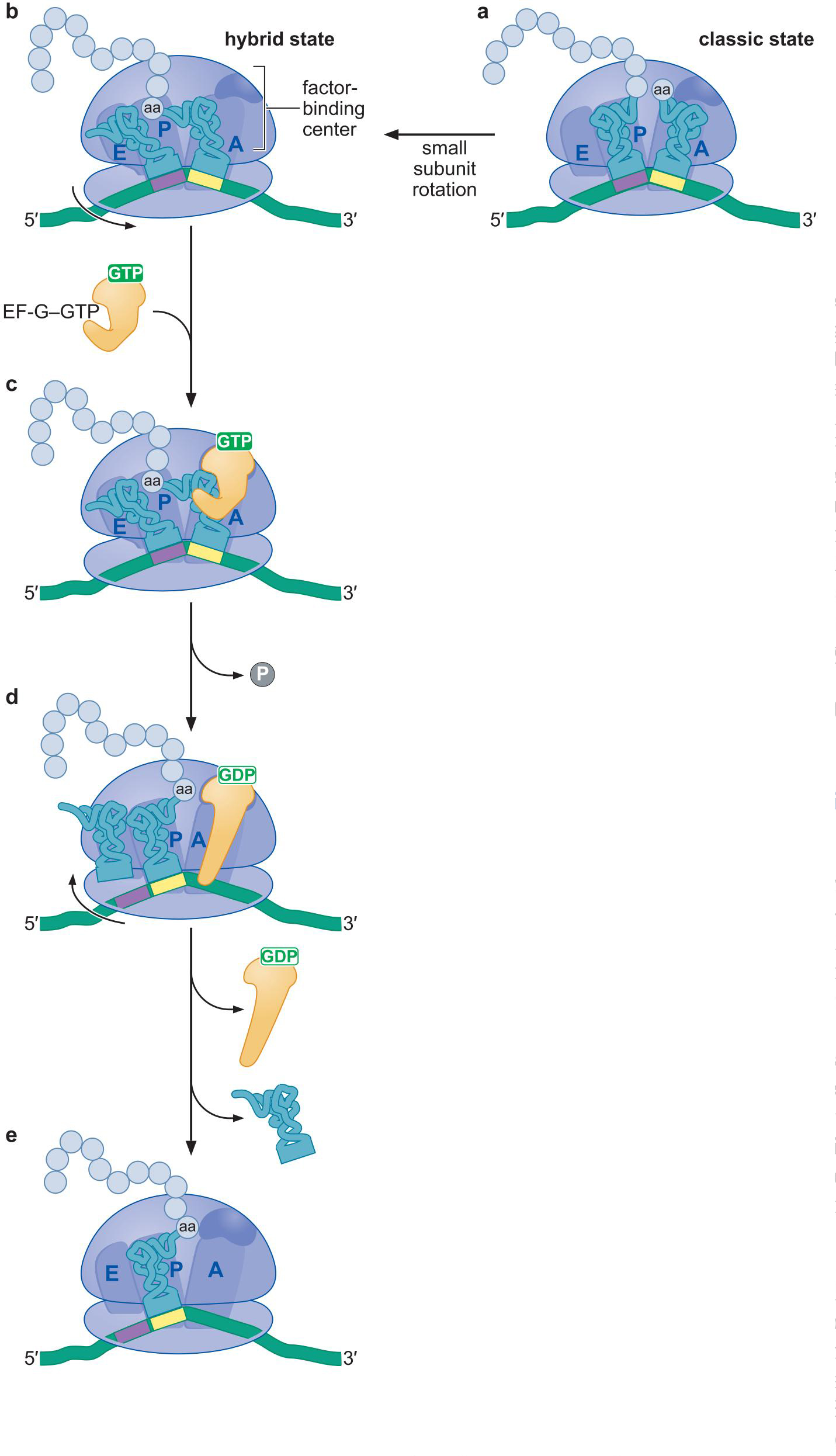
4、肽键的形成启动大亚基中的易位反应
- 大亚基的易位先于小亚基
5、EF-G推动易位
- EF-G-GTP结合在大亚基上
- EF-G-GTP水解形成EF-G-GDP 构象改变,占据A位点
- 推动小亚基中的易位
- 多米诺骨牌似的使得P位的tRNA易位向E
- 稀有移码tRNA(4bp)补偿移码错误
EF-G-GTP 的 分子拟态:
- EF-G-GTP 的形态很像 EF-Tu-GTP=tRNA复合物
6、EF-Tu-GDP 和 EF-G-GDP 在新一轮延伸前,要将GDP换成GTP
- EF-Ts促进 GDP与 EF-Tu解离
- EF-G 与 GDP亲和力<GTP亲和力,直接换

7、形成肽键的一个循环消耗两个GTP,一个ATP
- 氨酰-tRNA的形成 1ATP
- EF-G-GTP 牵引氨酰tRNA正确匹配 1GTP
- EF-G-GDP推动易位 1GTP
翻译的延伸中,原核与真核很相似
EF-Tu :eEF1
EF-G :eEF2
七、翻译的终止
1、释放因子在终止密码子的作用下终止翻译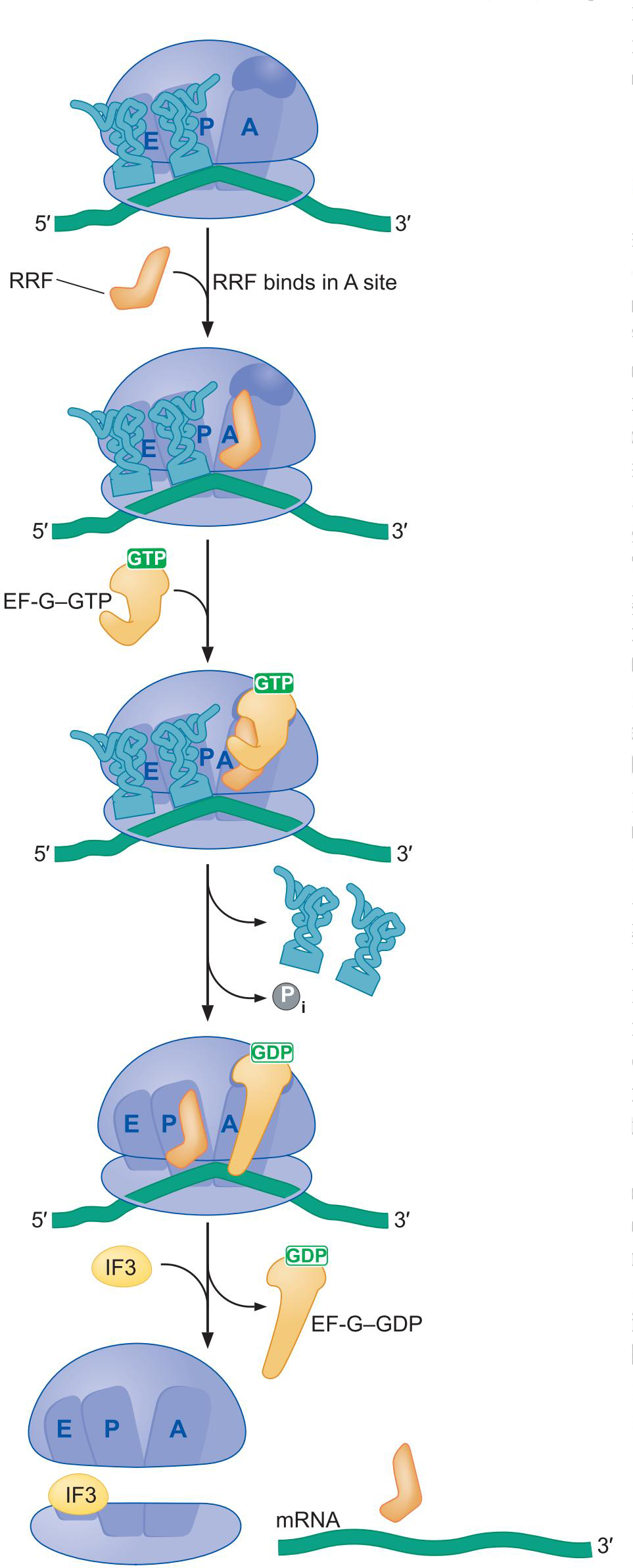
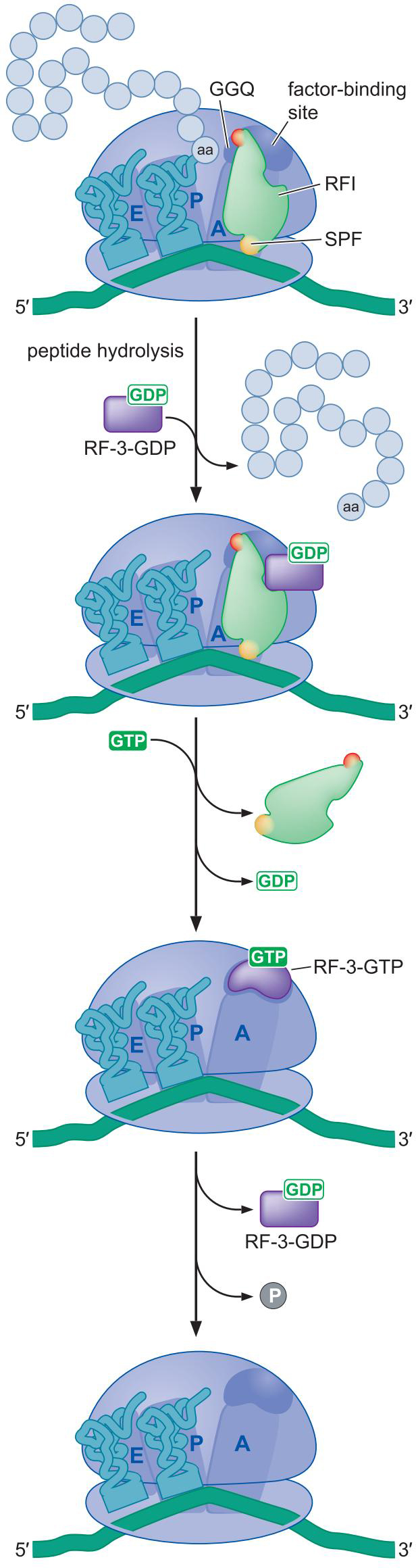
- 核糖体循环在 终止密码子进入 A 位点后,就停止
- 终止密码子被释放因子识别 release factor
- 两类释放因子
- I 类释放因子识别终止密码子:RF1 、RF2
- II 类释放因子:释放肽链后,刺激 I 类因子从核糖体解离
2、I 类因子的一小段区域识别终止密码子,并催化多肽链的释放
- I 类释放因子上的肽反密码子识别终止密码子
- I 类释放因子的GGQ序列促进了多肽链的释放
- I 类释放因子功能上模仿了tRNA
3、II 类释放因子的功能受GTP、GDP调控
4、核糖体循环因子模仿tRNA
- 核糖体循环因子(RRF)识别结合A位点
- EF-G-GTP结合核糖体,GTP水解,占据A位点,促使RRF易位至P
- 两个tRNA脱落
- IF3 加入 EF-G-GDP脱落
八、翻译的调控
- 翻译主要在起始阶段调控
1、蛋白质或RNA在RBS附近结合负调控细菌翻译的起始
- RNA分子的配对会被核糖体打破,后续的核糖体可以识别

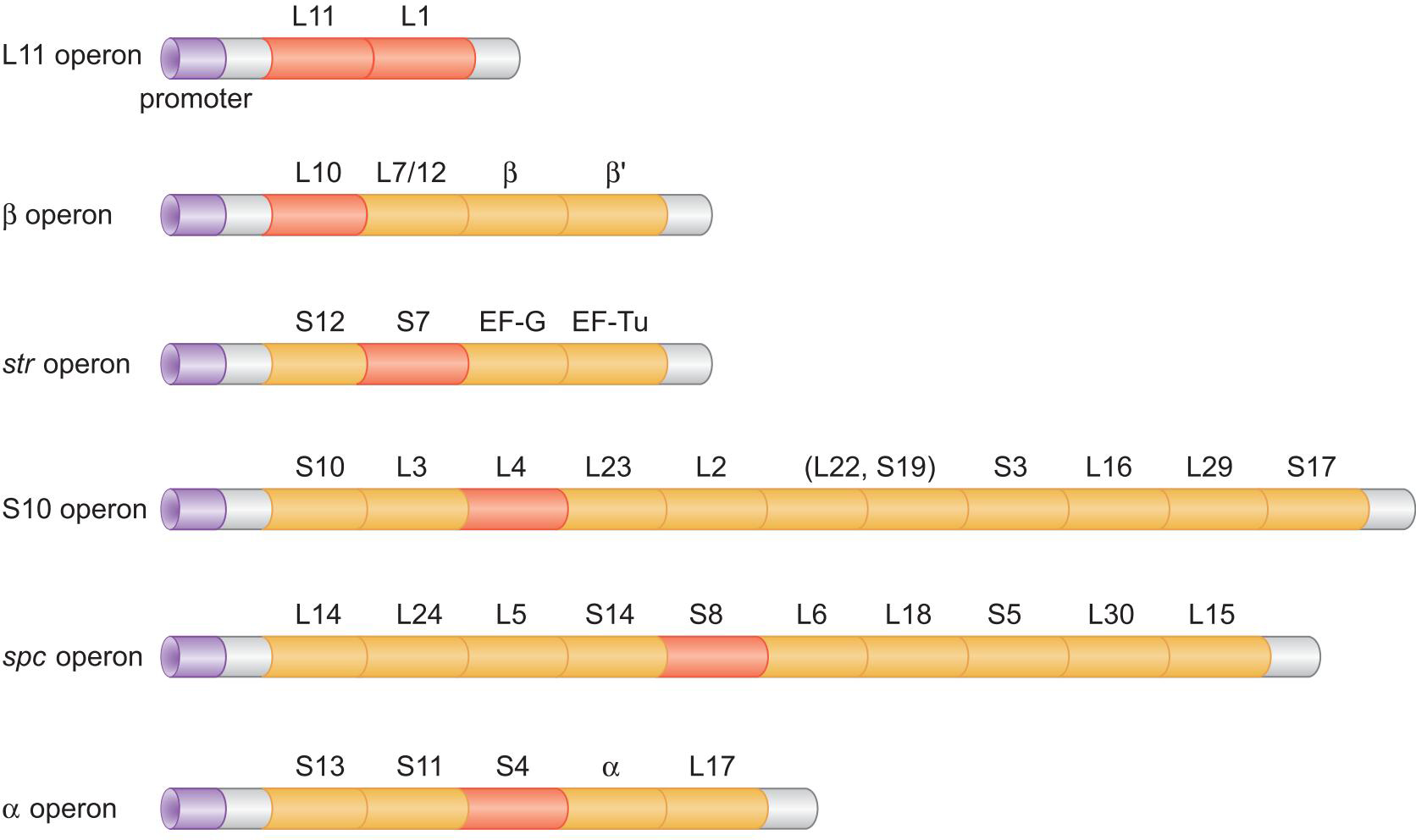
2、原核翻译调控:核糖体蛋白是其自身结合翻译的抑制因子
- 原核核糖体蛋白基因的协同调控通过若干操纵子得以简化
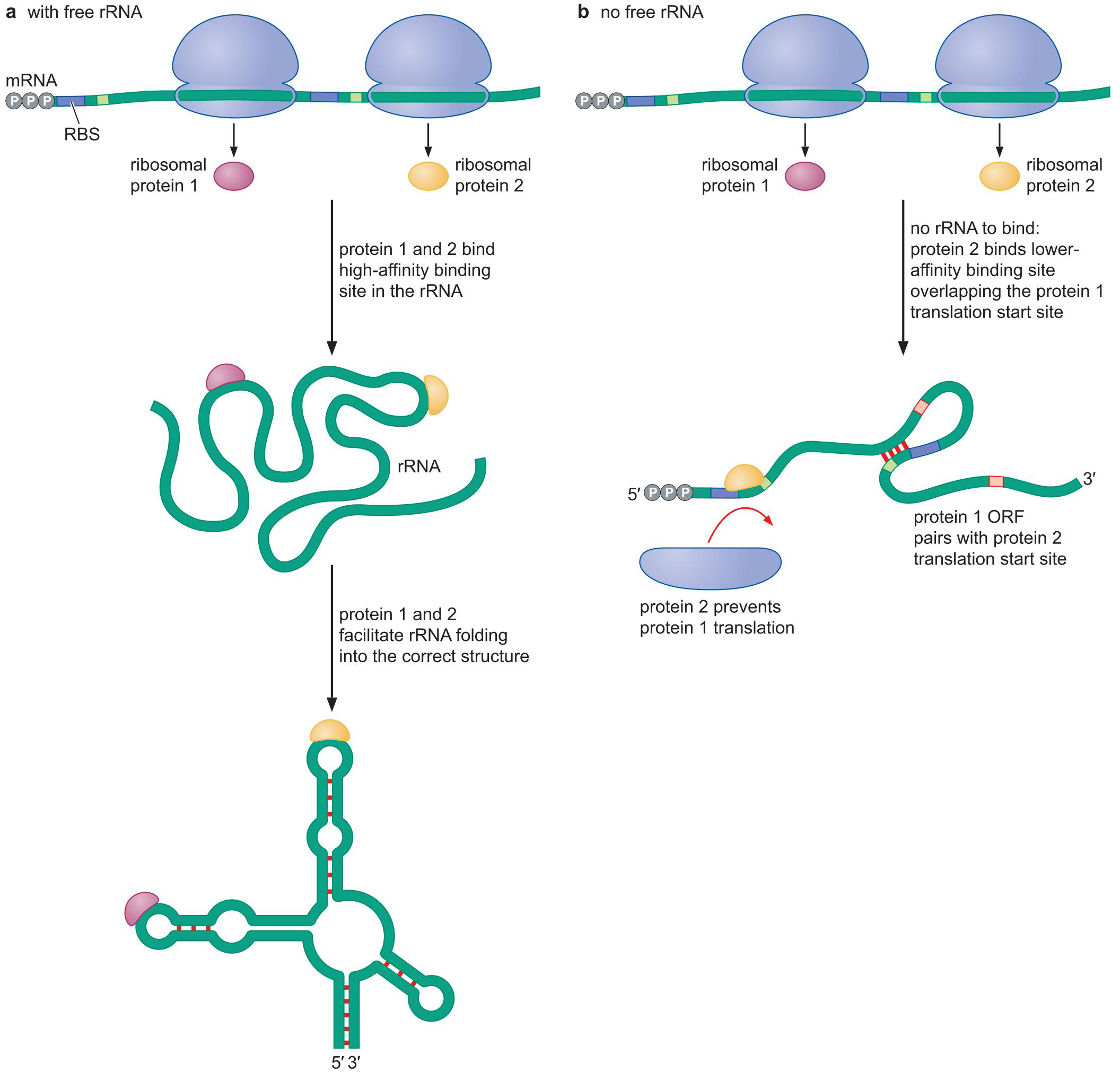
- 核糖体蛋白翻译的自我抑制:
- rRNA充足时,核糖体蛋白首先结合rRNA
- rRNA缺乏,核糖体蛋白结合自己的mRNA RBS附近
- 阻止翻译
3、真核翻译总体调节因子以mRNA识别及起始tRNA结合所需关键因子作为靶标
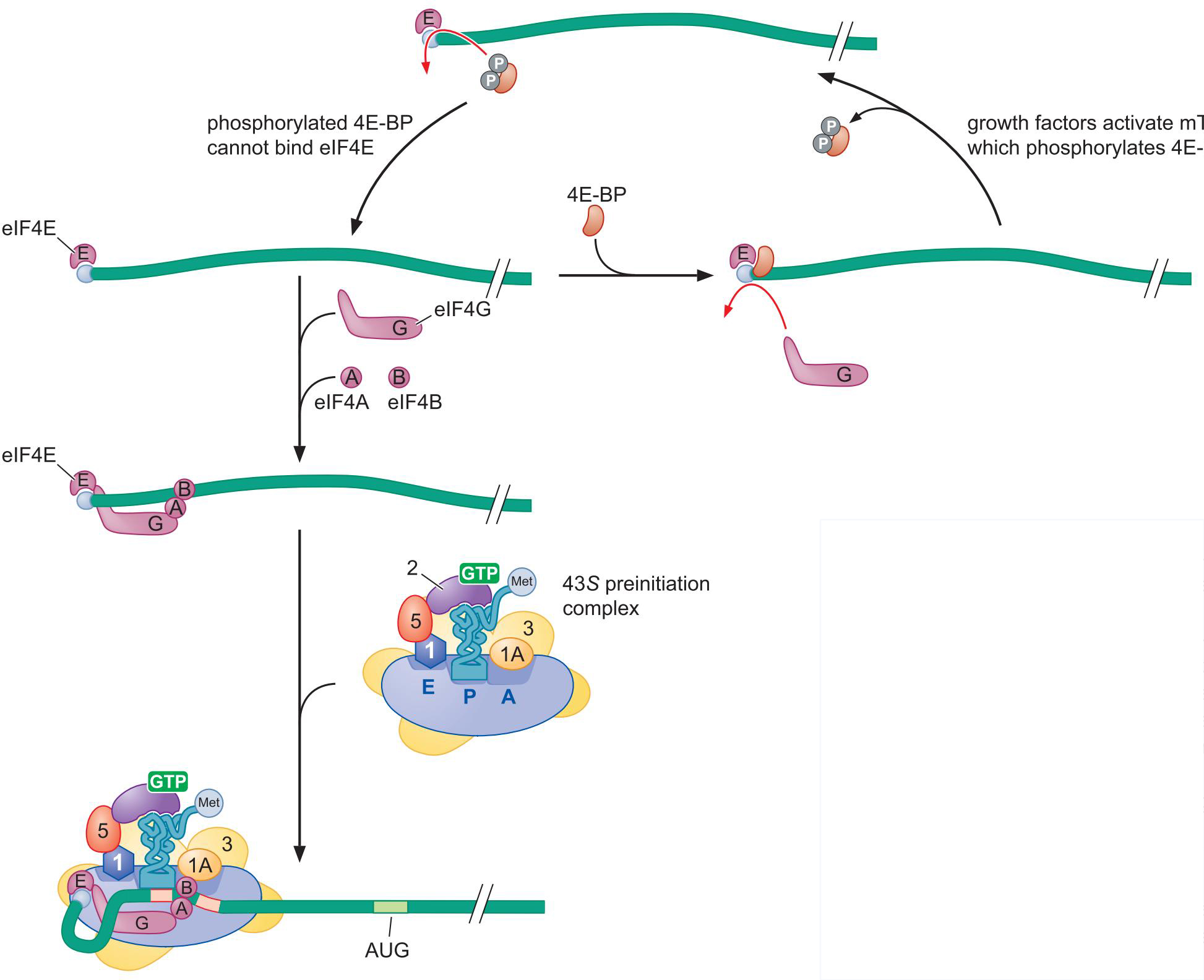
- mTor因子对4E-BP的磷酸化与翻译的调控
- 磷酸化的4E-BP不能结合5'Cap,翻译进行
- 这是化疗药物的靶点 雷帕霉素
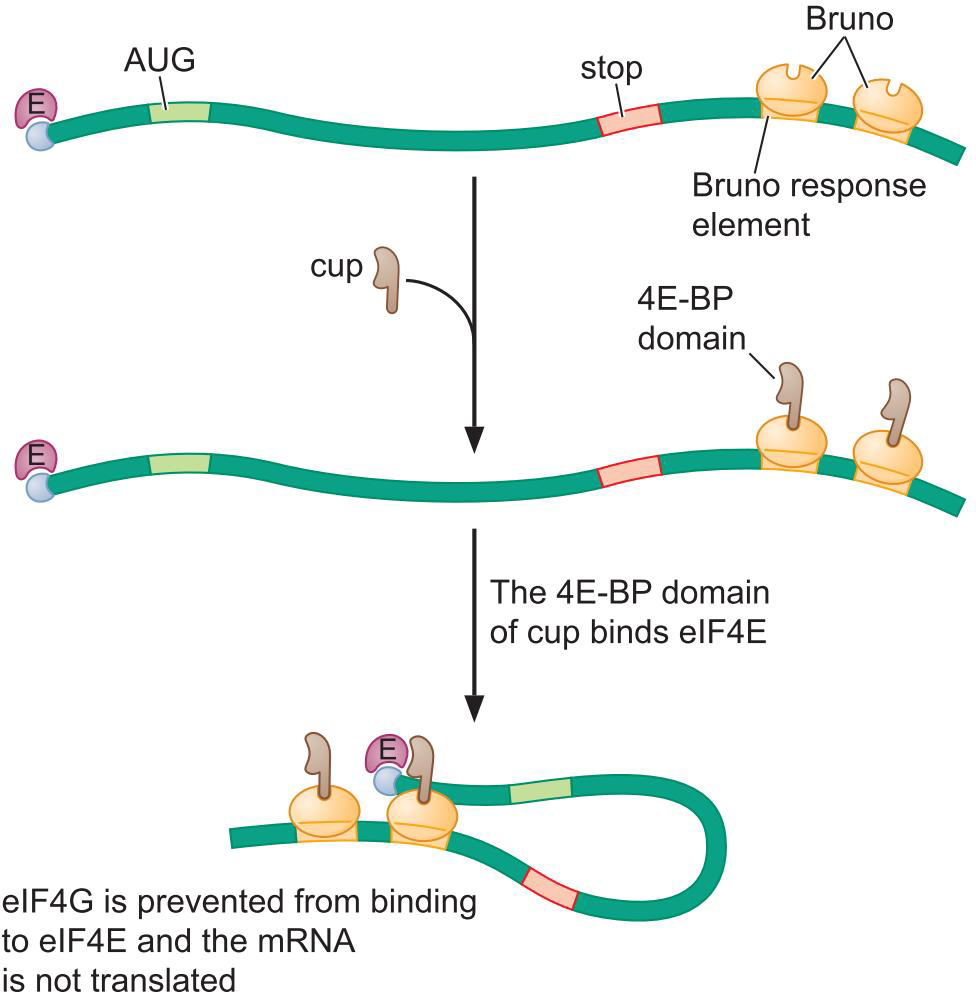
4、对mRNA特异的4E-BP进行翻译空间的调控
- 果蝇卵母细胞护送Oskar mRNA从前向后运输
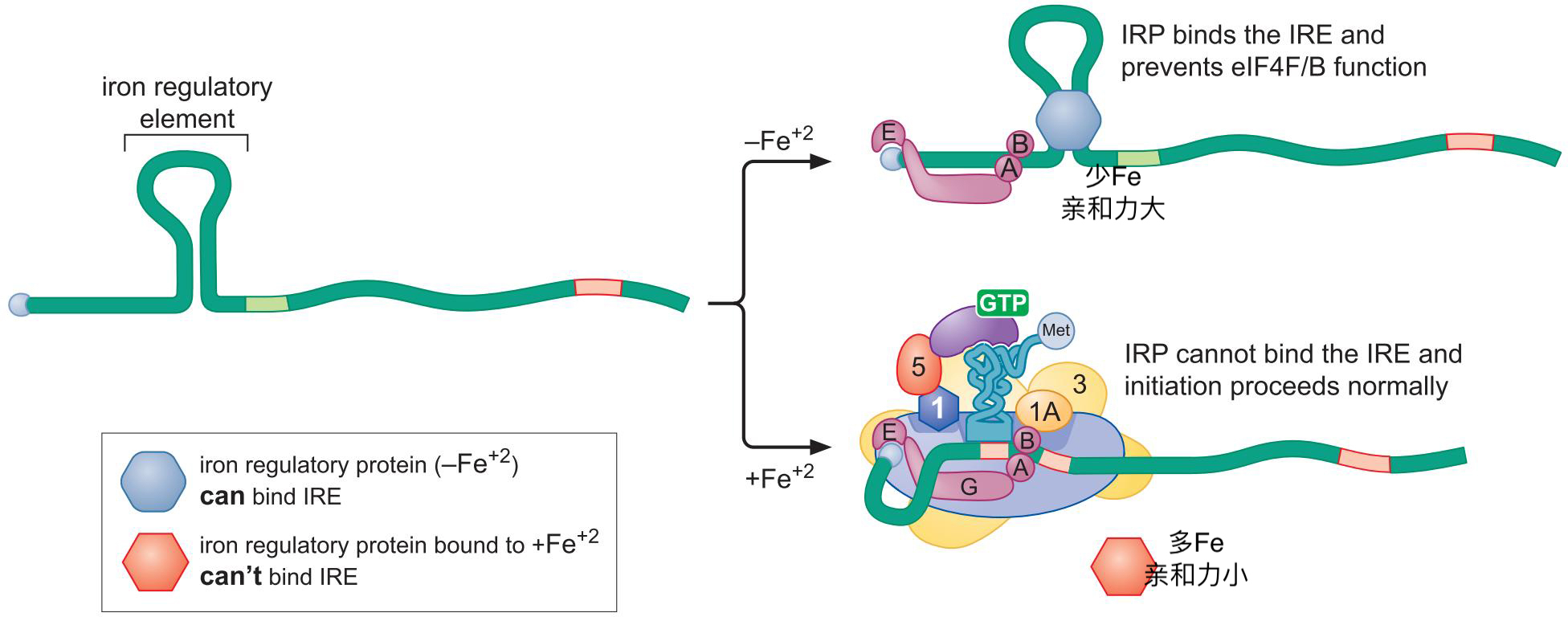
5、铁离子调控转铁蛋白翻译
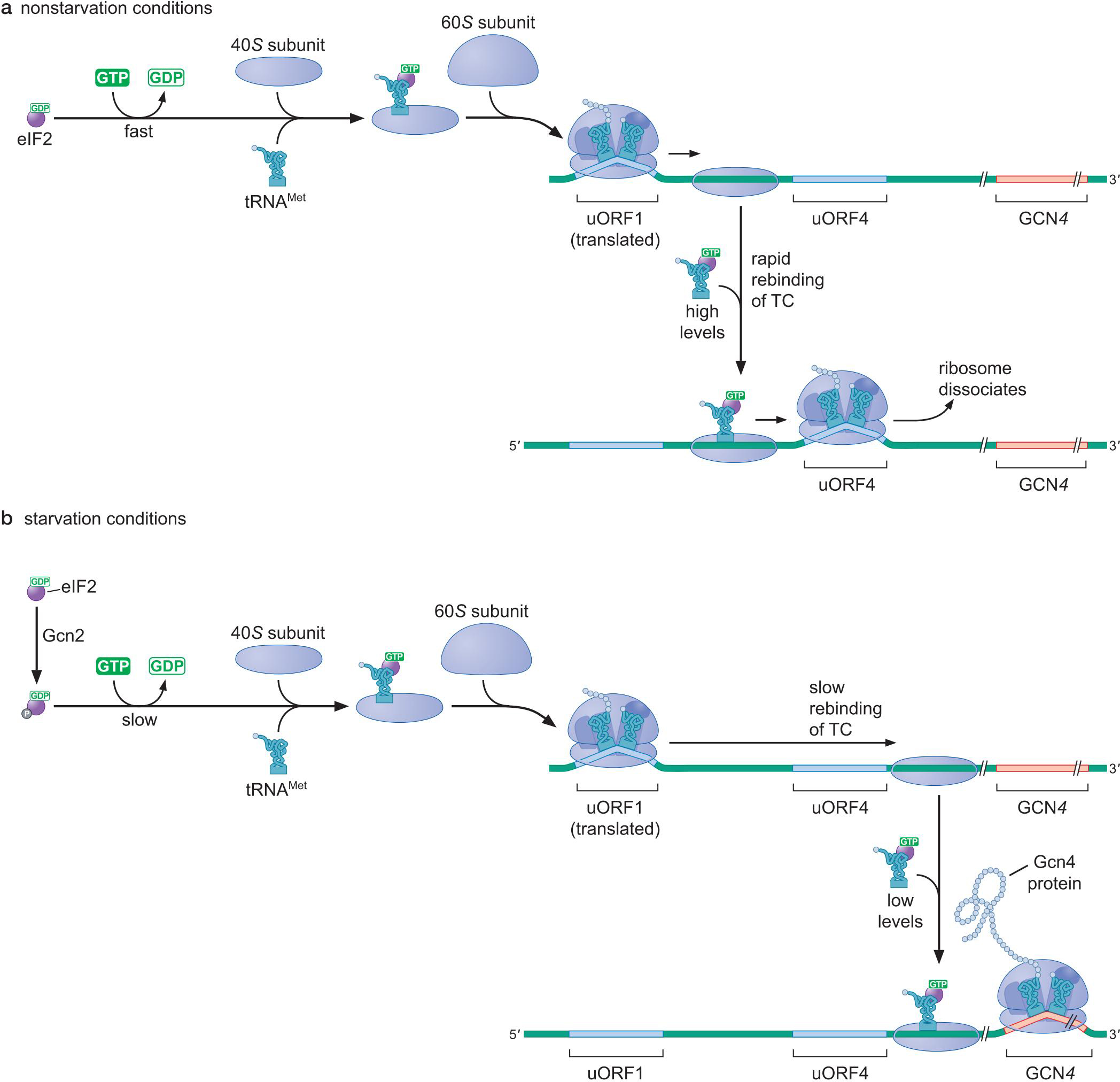
6、酵母转录激活因子Gcn4 的翻译受到上游短ORF及三元复合物丰度的调控
- Gcn基因上游4个小的可读框
- 翻译完第一个可读框的核糖体剩下一个小亚基在mRNA上
- 氨基酸充足,因素介导,小亚基迅速结合三元复合物,寻找下一个ORF起始密码
- 氨基酸缺乏时,小亚基结合三元复合物变慢,小亚基移出小ORF区后,才结合上,识别下游Gcn4基因起始密码,翻译
多聚核糖体谱
- 被核糖体结合多的就是翻译活跃的

九、依赖翻译的mRNA和蛋白质稳定性调节
- mRNA会出错,但在翻译中被识别出来之前,它是稳定的
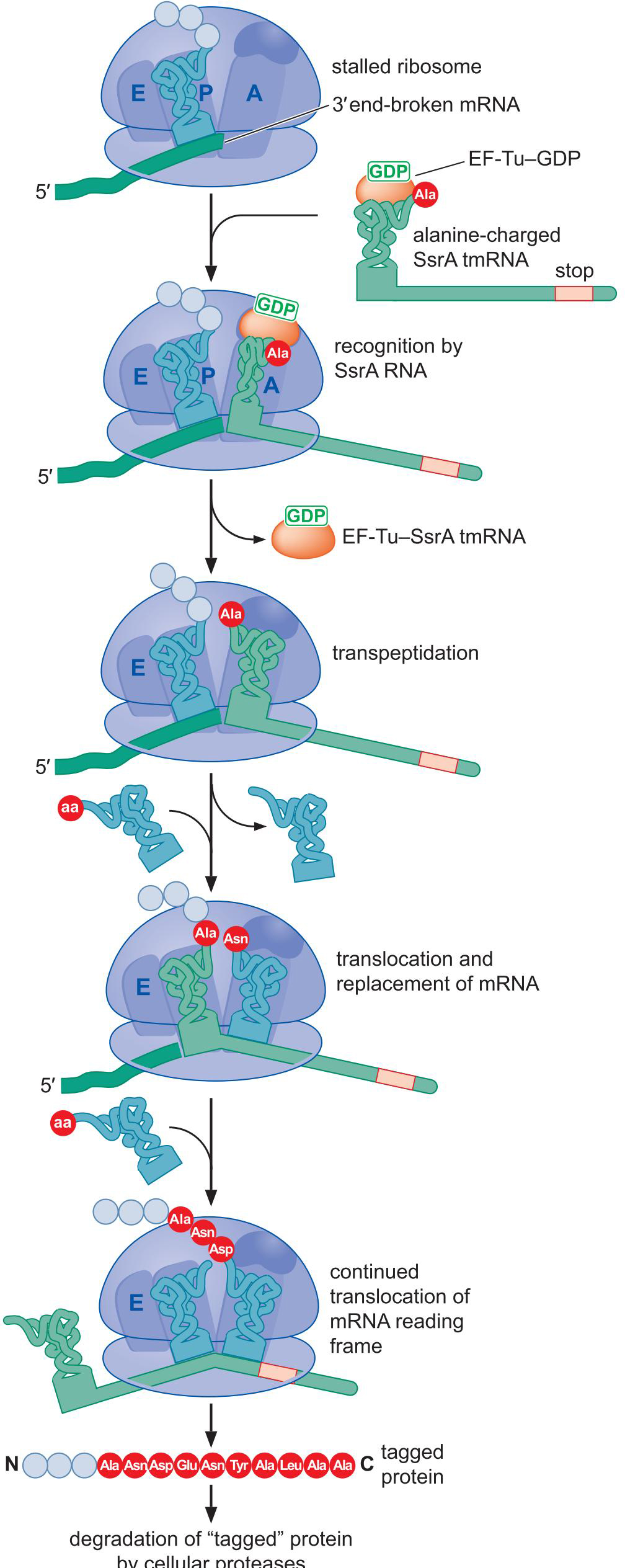
1、原核生物
- 缺乏终止密码子
- 核糖体在3'末端停滞,A位空缺
- SsrA RNA 加入A位点
- 类似正常的转肽和易位反应,将前面的tRNA和mRNA踢出
- SsrA RNA加入后翻译出来的多肽带有10个氨基酸的标签,招募蛋白将其降解
2、真核生物
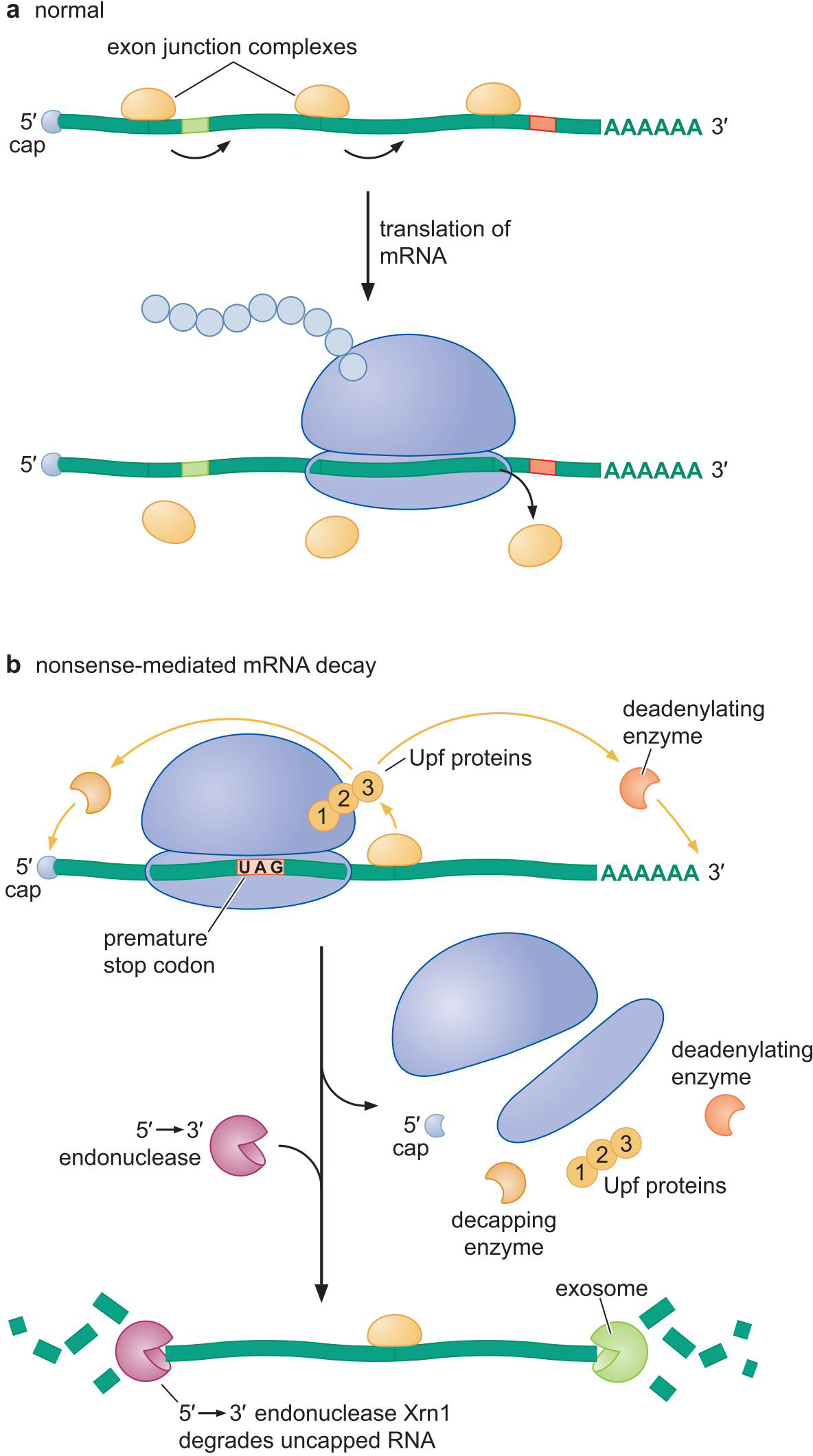
① 无意义密码子介导的mRNA衰减
- 真核生物mRNA经过剪接后,会留有 外显子拼接复合物
- 正常的翻译,核糖体会将一路上的 拼接复合物清除
- 如果提前终止了,就有 拼接复合物无法清除
- 招募5'->3'、3'->5'外切酶清除mRNA
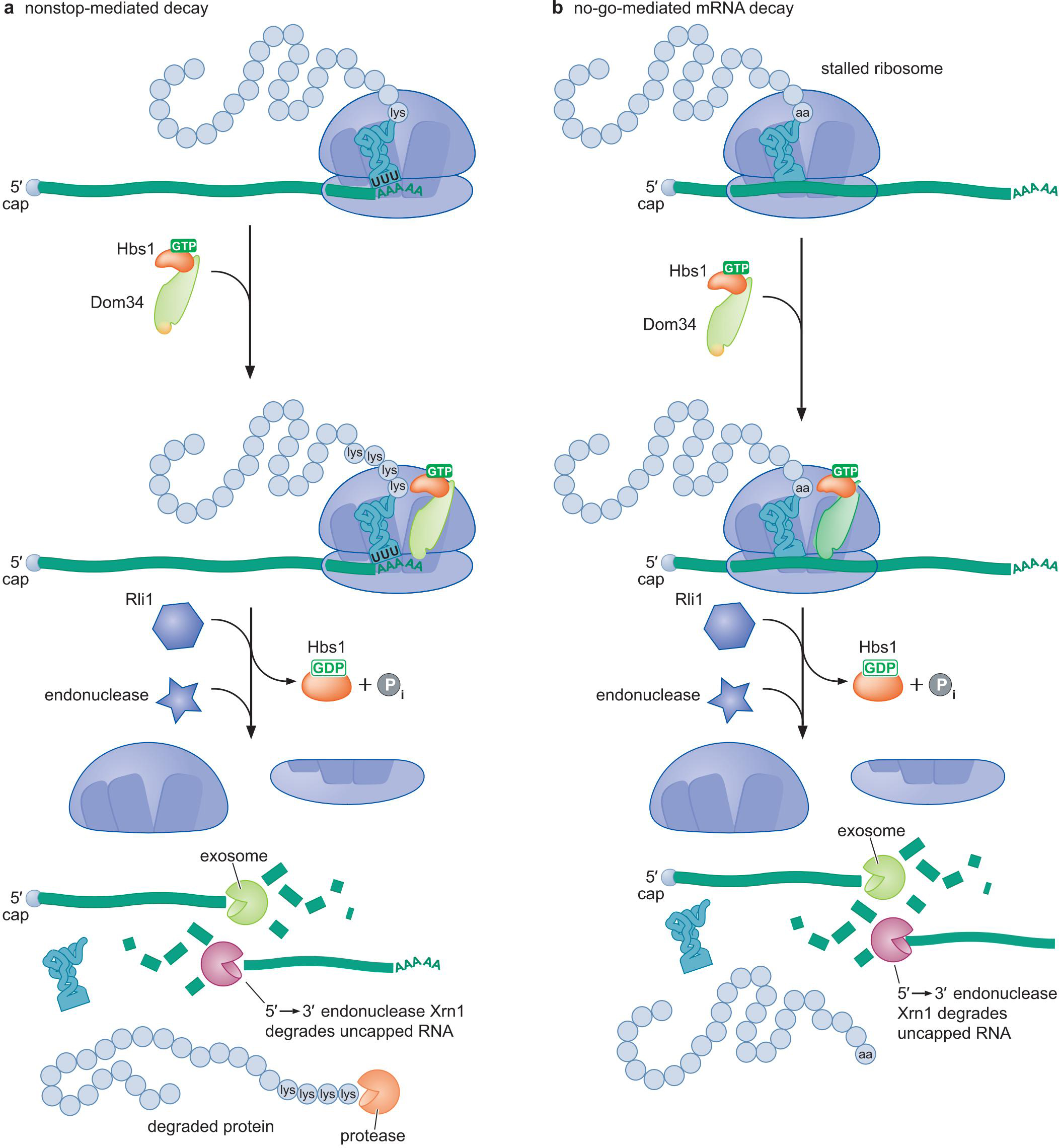
② 无终止密码子介导衰减
- 真核mRNA以poly-A结尾
- 无终止密码子,会翻译出尾端的多个 AAA(赖氨酸)
- 招募降解mRNA 和 蛋白质
③ 末端终止介导的衰减 no-go decay
- 当核糖体因为二级结构,或者相关tRNA不够用时,停滞,招募降解
十、遗传密码
1、遗传密码的简并性
- 密码子第一位改动改变氨基酸
- 第二位:嘧啶往往疏水;嘌呤往往极性
- 第三位:较少会发生氨基酸的改变
2、遗传密码的摆动性
|
反密码子 |
密码子 |
|
G |
U 、C |
|
C |
G |
|
A |
U |
|
U |
A、G |
|
I |
A、U、C |
3、遗传密码遵循3条规律
- 5'->3' 阅读
- 密码子不重叠,信息间不重叠
- 固定可读框上翻译
4、突变与抑制突变
- 错义突变
- 终止突变
- 移码突变
- 一个碱基缺失导致移码突变:可以由随后增加一个碱基的突变补偿
- 因为突变产生的提早的终止子可以被特殊的tRNA抢先读取,
5、遗传密码几乎通用
MIT molecular Biology 笔记10 翻译的更多相关文章
- MIT Molecular Biology 笔记6 转录的调控
视频 https://www.bilibili.com/video/av7973580?from=search&seid=16993146754254492690 教材 Molecular ...
- MIT Molecular Biology 笔记5 转录机制
视频 https://www.bilibili.com/video/av7973580?from=search&seid=16993146754254492690 教材 Molecular ...
- MIT molecular Biology 笔记11 位点特异性重组 和 DNA转座
位点特异性重组 和 DNA转座 视频 https://www.bilibili.com/video/av7973580/ 教材 Molecular biology of the gene 7th ed ...
- MIT molecular Biology 笔记8 RNA剪接
视频 https://www.bilibili.com/video/av7973580/ 教材 Molecular biology of the gene 7th edition J.D. Wat ...
- MIT Molecular Biology 笔记7 调控RNA
视频 https://www.bilibili.com/video/av7973580/ 教材 Molecular biology of the gene 7th edition J.D. Wat ...
- MIT Molecular Biology 笔记1 DNA的复制,染色体组装
视频 https://www.bilibili.com/video/av7973580?from=search&seid=16993146754254492690 教材 Molecular ...
- MIT Molecular Biology 笔记4 DNA相关实验
视频 https://www.bilibili.com/video/av7973580?from=search&seid=16993146754254492690 教材 Molecular ...
- MIT Molecular Biology 笔记3 DNA同源重组
视频 https://www.bilibili.com/video/av7973580?from=search&seid=16993146754254492690 教材 Molecular ...
- MIT Molecular Biology 笔记2 DNA的突变和修复
视频 https://www.bilibili.com/video/av7973580?from=search&seid=16993146754254492690 教材 Molecular ...
随机推荐
- Microsoft Visual C++ 14.0 is required. Get it with "Microsoft Visual C++ Build Tools"
https://blog.csdn.net/saucyj/article/details/79043443
- 正则表达式 re sys os random time 模块
今天学习内容如下: 1.正则表达式 百度正则表达式在线测试,可以练习 正则表达式本身也和python没有什么关系,就是匹配字符串内容的一种规则.官方定义:正则表达式是对字符串操作的一种逻辑公式,就是用 ...
- hdu 1598 (并查集加贪心) 速度与激情
题目传送门:http://acm.hdu.edu.cn/showproblem.php?pid=1598 一道带有贪心思想的并查集 所以说像二分,贪心这类基础的要掌握的很扎实才行. 用结构体数组储存公 ...
- grep如何进行正则表达式查找
字符类 字符类的搜索:如果我想要搜寻 test 或 taste 这两个单字时,可以发现到,其实她们有共通的 't?st' 存在-这个时候,我可以这样来搜寻: [root@www ~]# grep -n ...
- 如何在angularjs里面选择一个按钮而不改变其他按钮的颜色
var selectJson = { "background-color": "#FF0000", /* Green */ "border" ...
- Windows 下 Phpstrom 配置git使用
首先先去下载 git 下载链接 https://git-scm.com/download/winphpstrom 配置git 链接 http://jingyan.baidu.com/artic ...
- Delphi各种Socket组件的模式和模型
Delphi各种Socket组件的模式和模型 Delphi的大多数书籍里面都没有提到delphi的各种socket通信组件的模式和模型,有的书只讲解了windows的socket模式和模型,并没有归纳 ...
- Linux 内核态与用户态通信 netlink
参考资料: https://blog.csdn.net/zqixiao_09/article/details/77131283 https://www.cnblogs.com/lopnor/p/615 ...
- dubbo学习(zz)
dubbo学习 博客分类: 开源软件 Dubbo是阿里巴巴SOA服务化治理方案的核心框架,每天为2,000+个服务提供3,000,000,000+次访问量支持,并被广泛应用于阿里巴巴集团的各成员站 ...
- Homebrew -- mac 缺失包补充工具
https://brew.sh/index_zh-cn.htmlhttps://brew.sh/ 非root权限下运行 # github 源代码 https://github.com/Homebrew ...